コガネムシは円偏光(第3版1稿)


図:トルコ産のコガネムシ。左は左円偏光板、右は右円偏光板を通して観察したもの。
この読み物は昔別のサイトで公開していたものです。今から見ると書き直したいところもあり、サイトが使えなくなるタイミングで公開を停止したのですが、虫事に円偏光かをチェックしたデータなどは、興味のある方もいらっしゃるかと思い、あらためて公開します。
全面改訂版:「甲虫の構造色(コガネムシを中心に)に」もどる
第3版前書き
コガネムシの反射光が円偏光性を示すことに関しては、自分の中では完結した話になっていて、もはや改訂の必要はないと信じていた。ところが、2009年になって予想していなかったことが2つ生じた。最初の出来事はScience誌掲載されたコガネムシの表皮構造の論文で、コレステリック液晶の欠陥に類似した構造の存在を報告していた。二番目は、構造色研究会のシンポジウムでコガネムシと液晶の関係についての話をという依頼が舞い込んできたことである。折角に機会なので、コガネムシの表皮を自分でも少しばかり顕微鏡をのぞいてシンポジウムの資料を準備した。それを表に出さずにおくのも勿体ないので、その時の資料を含めてこのWebを改訂することにした。前の版との違いは、表皮構造に関する部分なので、前の版を読まれている方は、そこだけ目を通せば良いと思う。構造色
昆虫の中には、鮮やかな色彩を示すものも多い。どうして、そんな色彩になったのかは生物学的な問題であるが、どのような機構で色彩を示すかとなると、物理や化学の問題となる。実際、20世紀の初めのころには、南米に生息するモルフォ蝶の発色機構を巡って、色素派と構造発色派が議論を行っていた。色素派とは文字通りモルフォ蝶の発色の原因を羽根に含まれているであろう青色域に強い吸収を有する色素に求める一派である。青色域に吸収を持つと青く輝くというと変に聞こえるかもしれないが、吸収が強いと反射も強くなり金属光沢のような輝きを示すのは(吸収機構によらず)一般的な話である。例えば、金属類でも可視領域までもカバーする強い吸収が金属光沢の原因になっている。一方の構造発色派とは、モルフォ蝶の羽根の色は青色の色素によってではなく、青色の光を干渉で反射するような周期構造によるものであるとする一派である。
分析手段が進んだ今日にあっては発色の原因を突き止めることは容易である。モルフォ蝶に関しても、電子顕微鏡観察により光の波長程度の構造があり、それが発色の原因であることが明らかにされている。(モルフォ蝶の発色機構の詳細を知りたい方は「構造色研究会」のHPからコンテンツをお探し頂きたい。)。しかし、電子顕微鏡の無かった当時には、羽根をすりつぶして反射光が無くなる(色素だったら、すりつぶす程度では分子は破壊されないので色は残るはずである)といった、間接的な実験から2つの流派の間で正しさを巡る議論が行われていた。その議論の詳細を調べたことはないのだけれども、こうした昔話を聞くと、装置がなかった分だけ工夫により仮説の検証を行おうとする迫力を感じ、逆に今日の科学研究の多くが工夫のないものに思えることがある。
閑話休題。甲虫類でも玉虫やコガネムシなどの中に構造発色を示すものがある。甲虫の構造色の発現機構に関しては、2009年になって、A. E. Seagoらのレビューが出されており[1]、その中では3種類+αに分類されている。彼らの分類に従うと、一番目はMultilayer Reflectorsで、2種類の屈折率の異なる光の波長程度の厚さをもった膜が積層しているものである(誘電体多層膜タイプ)。二番目は3-D Photonic Crystals タイプで、微小球が周期的に配列したような構造になっているものである。三番目はDiffraction Gratingsで、積層軸が体表面に垂直方向にある積層構造により回折格子効果がでるものである。そのほかとしては、散乱構造による白色なども構造色としてとりまとめられている。
この論文の中では、円偏光を反射するものはCircularly polarizing reflectorsとして、Multilayer Reflectorsの一種として取り扱われており、birefringent chitin layers are deposited helicallyと説明されている。しかしながら、液晶をやっている立場からするとlayersという言葉に引っかかりがある。なぜ、引っかかりがあるのかを続いて説明することにしよう。コレステリック液晶
コレステリック液晶は液晶の一種である。歴史的にコレステロールの誘導体で始めて観測された状態で[2]、特異的な光学特性を示すのでコレステリック液晶と呼ばれるようになった。液晶の分類として、よくネマチック、コレステリック、スメクチックという3種類が上げられるが、コレステリック液晶は純粋に学問的にはネマチック液晶の一種で、分子が不斉構造を有する(あるいは不斉構造を持つ分子が混ぜられている)ものである。ネマチック液晶は液体と結晶の中間状態である。一般に液晶分子は細長い形状をしている。このような分子は結晶状態では、重心位置の長距離秩序の他にも、分子の方向の長距離秩序を持っている。一方、液体状態においては、重心位置、分子の方向とも秩序を失いランダムな状態となる。通常の物質では、重心位置と分子の方向の長距離秩序は融点で同時に消滅する。しかし、分子によっては、昇温により重心位置の秩序性が先に失われ、結晶のように方向の揃った分子が液体のように重心位置の秩序なく流動性を保つような結晶と液体の中間状態が出現する。これが、ネマチック液晶である。ネマチック液晶をさらに昇温すると、分子の方向の秩序も失われ、最終的に普通の液体となる。ネマチック液晶が重心の長距離秩序を持たないということは、層ということばとは無縁の存在であることを示している。というのは、層というのは一般に明示的に区別できる何らかの厚みをもった存在を意味するのだけれど、そのような平面的なシートをネマチック液晶の中に見いだすことは出来ないからである。そして、事情はコレステリック液晶でも同じで、分子の平均的な方向は螺旋を描いて変化していくが、それは、連続的であって、そこに層を見いだすことは出来ないのである。
 図:液晶を構成する分子の一つ(上)と液晶の集合状態(下)。分子の長軸方向は大体揃っている。
図:液晶を構成する分子の一つ(上)と液晶の集合状態(下)。分子の長軸方向は大体揃っている。
 図:コレステリック液晶の構造。なかなかうまく描けない…。液晶分子は常に画面奥方向への軸に垂直である。手前では上を向いている分子が真中では横を向き、奥ではまた上を向いている。
図:コレステリック液晶の構造。なかなかうまく描けない…。液晶分子は常に画面奥方向への軸に垂直である。手前では上を向いている分子が真中では横を向き、奥ではまた上を向いている。
上に記したように分子が不斉構造(不斉中心)を持っていると、分子長軸の配向方向が空間的に変化する。分子長軸の向きが一回転する長さをラセンピッチと言う。上の図ではラセンピッチの半分を描いている。コレステリック液晶では、この周期構造による反射が生じる。反射光の波長はラセンピッチを液晶の屈折率で割った値(光学的なラセンピッチ)となる。そしてまた、らせんのねじれ方向と同じねじれの光のみ反射する。それ故に、コレステリック液晶の反射光は円偏光となる。光学的ラセンピッチが可視領域にあれば、可視領域に反射が生じる。ラセンピッチは温度により変化する。これを利用したのが液晶温度計である。
ある種のコガネムシが円偏光を反射することは、それらの羽にはコレステリック液晶類似構造が存在することを示唆している。もちろん、コガネムシの羽は固体であるのでコレステリック液晶ではなく、それが固定したラセン構造というべきものであろう。人工の高分子コレステリック液晶の中にも急冷によりラセン構造が固定され鮮やかな反射光を示す高分子フィルムとなるものもある。
いくつかのコレステリックタイプの虫の写真
ここで、コレステリックタイプの虫の写真をお目にかけよう。これからお見せする写真は、いずれも左側が左円偏光板を、右側が右円偏光板を通して撮影されたものである。なお、円偏光が何か判らず知りたい方は付録に簡単な説明があるのでそれをまずご覧頂きたい。
まず最初はページの初めにも示したトルコ産のコガネムシである。
左円偏光では色彩が見えて、右円偏光では黒くなることから、この虫のラセン構造は左ラセンであることが分かる。緑色の部分も赤色の部分も同じく黒くなることから、緑の部分も赤の部分もラセン構造による構造色である。これはそれぞれの部分でラセンのピッチが異なることを示唆している。写真では差はそれほどはっきりしないが前足の部分に着目すると左の方は青色系の発色をしており、右側の方は黒くなっている。足の部分も構造色により発色しており、その色調は胴体部分と異なっていることが分かる。
右円偏光板を通した画像は黒くなっているが、これは、このコガネムシが羽の下側に黒色色素層を持っているためである。コレステリックタイプの場合には、構造色による反射率の最大値は、発色している波長域でもある例外を除いて50%となる。もし、左円偏光が羽を通過後に反射されて戻ってくると、コガネムシは全体にもっと明るくなり淡い色調になる。
次はエクアドル産のものである


この虫も左ラセンである。
外国の虫が続いたので、つぎに国内の虫をお見せしよう。
![]()
![]()
これは、鹿児島の沖合の島のコガネムシである。この虫も左円偏光のみ反射しており左ラセンになっている。右円偏光でみると、色素の欠落した斑入りになっている。左円偏光の写真でも、その部分は同じような色調になっている。この部分がラセン構造も存在していないのか、それともラセン構造はあるけれどコントラストが悪くて他との違いがはっきりしているのかは、確認していない(そういう目できちんとルーペで拡大して見ていなかった)。この虫以外にも、そこらへんに結構みかけるドウガネブイブイもコレステリックタイプの虫で、左ラセンを持っている。
つづいては、コガネムシの中のコガネムシ、ダイコクコガネの一種である。


この虫も左ラセン構造となっている。これまでのコガネムシと少しばかり違った印象の外観であるけれど、コレステリックタイプという点では共通の要素を持っているのである。この虫を最初に見たのは、箱根にある「生命の星地球博物館」でニジイロダイコクコガネがかなりたくさん展示してある。トルコのコガネムシを見る前だったので、一つの虫の中で構造色により違った色に発色した部分があるのが非常に不思議で新鮮に思えて印象に残っていた虫である。
続いては、宝石のような虫、その名もプラチナコガネの一種である。この手の虫の反射光が円偏光であることは、実に20世紀初頭には確認されていた[3]。


これは、南米に生息する虫でこれはコスタリカ産のベーツプラチナコガネという虫である。この虫も左ラセンタイプである。これまでの虫が緑とか赤とか、可視領域の光の一部を反射していたのに対して、この虫は少し青みを帯びているので、長波長側の反射率が少し低いようだけれど、可視領域ほぼ全体に渡って光を反射している。これは、実に驚異である。何しろ、コレステリック液晶の反射帯の幅は、液晶を構成する分子の長軸方向と短軸方向の屈折率の差に依存しており、屈折率差が大きいほど反射帯も広くなるのであるが、約380nmから800nmの可視領域全体が反射帯となるためには、屈折率が倍程度違わないといけないのである。通常の有機物の屈折率は1.5程度である。屈折率が小さい方の屈折率の値が1.5よりも小さいとしても1より小さいことはあり得ない(何しろ、反対側の光に対しては透明なのだ。)。すると、大きい方の屈折率は2以上となり、屈折率差(複屈折)は1以上あることになる。合成された液晶で屈折率差が大きいものでも、その値は0.4とか0.5がせいぜいである。それからすると1以上の屈折率差は考えられないほど大きな値である。そのような有機分子が見つかったら、かなり大きなインパクトがある。
残念ながら、この虫の羽にはそんな大きな複屈折を持つ物質は含まれていない。その換わりにラセンの周期が羽の厚さ方向で変化している。つまり、羽のある部分は青色の光を反射し、続いての部分は緑、そしてその先は赤という具合になっていて、全体として可視領域全部の光を反射しているのである。ベーツプラチナコガネについては分析はされていないが、同じくプラチナコガネ属に含まれるオプティマにおいて羽の斜め切断面の電子顕微鏡観察からラセン構造が分布を持っていることが明らかにされている[4]。
もっとも、金属光沢を持つプラチナコガネの秘密はそれだけではない。参照文献4には、これらのプラチナコガネは羽に尿酸分子を多く含んでいることも記されている。
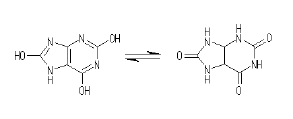
尿酸は上の図に示したような分子で、羽の他の構成分子に比べて大きな複屈折を示す。先ほど、プラチナコガネの複屈折は非常に大きいものではないと記したけれど、たとえラセンのピッチに分布があるとしても複屈折が大きい方が金属光沢を出すのに有利になる。これは、複屈折が大きい方が反射帯の幅が広くなるので、より少ないピッチのバリエーションで可視領域をおおえるようになることと、複屈折が大きい方が、より少ないラセンの巻き数で反射率が高くなるからである。参照文献4によると、これらのコガネムシをアルカリ溶液で洗って尿酸を溶かし出してしまうと羽の金属光沢が失われるそうである。
ところで、ベーツでは右円偏光での写真がそれほど黒くないことに気がついた方もいると思う。ベーツプラチナコガネでは報告されていないが、オプティマでは羽の裏側に黒色の色素を持っていないことが知られている。後で述べるように黒色色素の存在は鮮やかな発色をもたらす。可視領域全体で反射して特定の発色を示さない場合には、黒色色素の存在は必ずしも必要ない(ただし、これは人間の目から見てである。紫外や赤外に感度を持つ目を持つものからすれば違いがでるはずである。従って、黒色色素を持たないことが生物学的に意味があるのかはよく分からない。)。
最後に、例外的な虫をお目にかけよう。


この虫も南米コスタリカ産のプラチナコガネの一種、レスプレンデンスである。これまでと同様に左ラセンタイプである。と記すと、「そんなこと言ったって右側の写真がこれまでみたいに暗くないじゃないか」という声がとんで来そうである。でも足の部分をよく見て欲しい。左の写真では明るく写っているのに、右の写真では黒くなっているであろう。
このコガネムシの鞘翅の構造についても参照文献4に記載がある。それによると、レスプレンデンスもコレステリック構造を持っているが、2層の左ラセンの間にλ/2波長板相当の複屈折層を持っている。円偏光がλ/2板を通ると符号が反転する。つまり右円偏光は左円偏光に、左円偏光は右円偏光になる。外から無偏光が入ってくると外側の層で左円偏光が反射され、右円偏光は透過する。そして、右円偏光がλ/2板部分で左円偏光に変換され、続いて存在する2番目の左ラセン層で反射される。反射された左円偏光は再びλ/2板を通る時に右円偏光になり右円偏光として外に出て行く。こうして、この虫では見かけ上左円偏光も反射し、右円偏光も反射する。この結果、この虫は高い反射率を示す。下の図に幾つかのコガネムシの反射スペクトル測定結果を示すが、この虫のみ最大反射率が50%を越えて80%近くに達している。虫が金色になるのは、ベーツプラチナコガネと同様にラセン周期に分布があるためである。ただし、1層目と2層目で分布に違いがあるために、左円偏光板を通した時と右円偏光板を通した時では微妙に色合いが異なる。左の時は少し緑がかっており、右の場合にはオレンジがかっている。なお、足の部分はλ/2板を持たないので右円偏光では黒くなるのである。
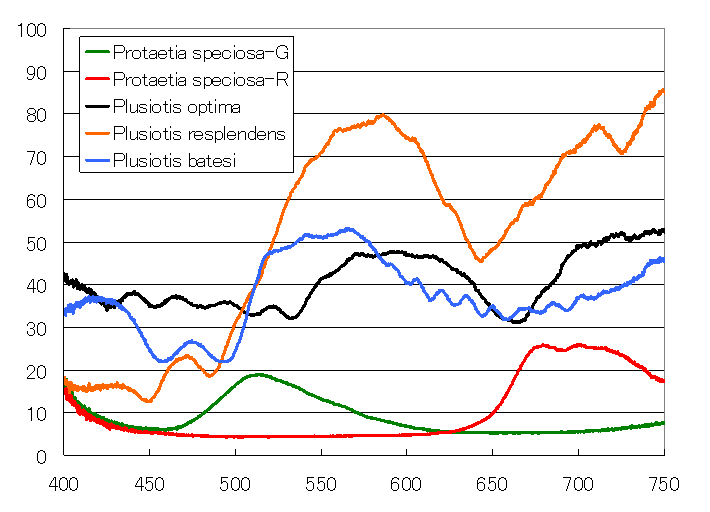
金属光沢を持つプラチナコガネは何種類もいるが、その中でも両方の円偏光をはねるのは私が見た限りではレスプレンデンスだけである。そういう意味で、まさに例外中の例外と言って良い。幸いなことにレスプレンデンスはプラチナコガネの中では比較的安価な部類に属する。この特異性故に人気が出てしまわないことを祈っている。
ここまでで示した例では全ての虫は左ラセンの構造を持っている。ただし、このことが、生物界における右と左の問題と必ずしも直接には結びつかないことは注意しておいた方がよい。というのは、確かに同じ分子からなるコレステリック液晶ではラセンの掌性は分子の掌性と1対1の関係があり、例えばL体分子が左巻ラセンを形成するなら、D体分子なら右巻ラセンを形成する。しかし、異なる構造を持つ分子間ではそうは行かない。同じL体から構成される分子からなるコレステリック液晶でも右巻ラセンもあれば左巻ラセンもある。例えば、コレステロールの誘導体の液晶は、側鎖に何をつけるかによってラセンの向きが異なることが知られているのだ。ここで紹介した甲虫の甲を形成する天然高分子が同じ物だったら、ラセンが同じ向きなのは必然であるのだけれど、化学的な違いがあったりしたら、同じラセンであることは、かなり偶然な出来事である可能性もある。
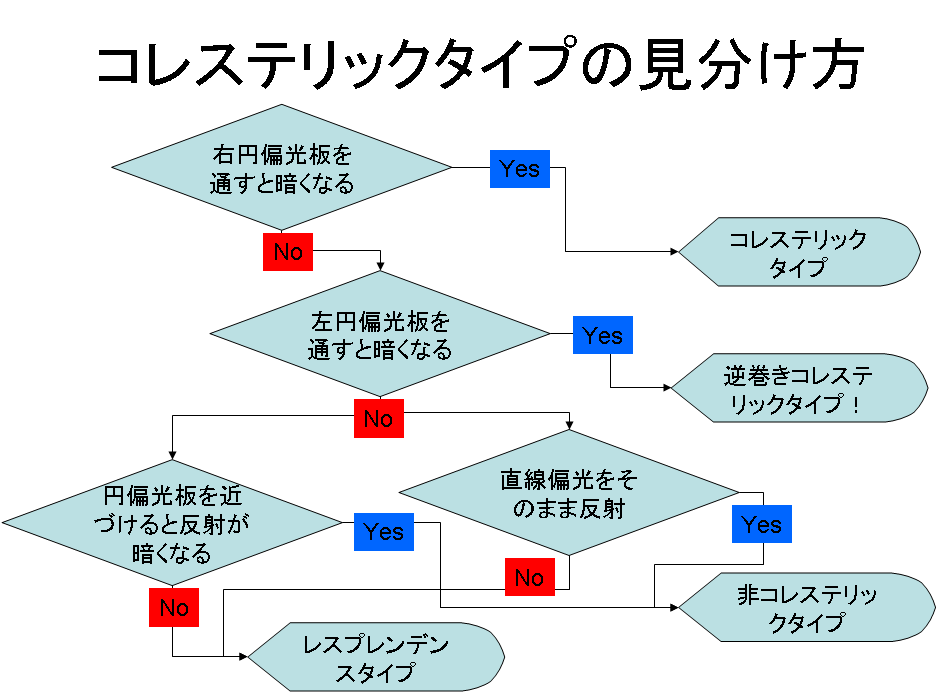
コレステリックタイプの昆虫の見分け方
コレステリックタイプのコガネムシはレスプレンデンスを除いては右円偏光板を通すと色を失ったり黒くなったりするので、右円偏光板を持ち歩いていて、それを通してみて色が失われれば、コレステリックタイプの昆虫であると結論できる。残念ながら変化が見られない場合は続いて左円偏光板を通して観察してみる。というのは、僅かではあるが、左円偏光板で変化が生じるコガネムシの報告があるのである。左円偏光を反射するコガネムシについては、突然変異で存在するという記述と、レスプレンデンスタイプのものを見誤ったのではないかとの指摘がある[5]。真偽は現在では明らかになっていないが、何れにせよ、0ではない可能性として頭に入れておく必要がある。
右・左円偏光板を通しても格段の変化が見られない場合には非コレステリックタイプである可能性が高い。とは言え、例外にあたっていない保証はないので、そのような場合には続けて、円偏光板を虫の鞘翅に密着させて反射を観察する。通常の誘電体界面では円偏光は反射時に符号が反転する。このため円偏光板を密着させると鞘翅からの反射光は現象する。一方、コレステリック液晶では、円偏光が符号を反転せずに戻ってくるので、密着させてもあまり暗くならない。もう一つの確認方法は、リングクロス偏光フィルターを使った撮影を行ってみた。これは、リングフラッシュに装着するアクセサリーで、発光部分とカメラレンズの前の部分で偏光方向が90度異なっている。このため反射時に偏光方向が変わらなければ光はレンズに到達せずその部分の反射は写らなくなる。一方、コレステリックタイプのコガネムシでは直線偏光を入射しても円偏光が戻ってくるのでカメラの前の偏光フィルターを透過する成分がある。図にチェックのフローチャートを示す。
図に、円偏光板を密着させた場合の様子を示す。左のコレステリックタイプでは、反射が確認されるが、右側の非コレステリックタイプでは、全体に暗くなってしまう。

続いてリングクロスPolフィルターを通しての違いをお目にかけよう。フィルタ自体は


である。左の非コレステリックは反射がなく黒くしか写らなくなる。
コガネムシ鞘翅表面の構造
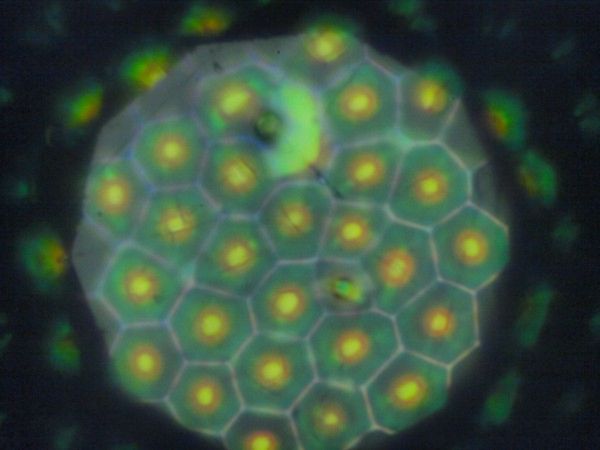
2009年にScienceにコガネムシに関する論文が掲載された[6]。その論文によると、コガネムシの表皮には、コレステリック液晶が空気界面で自発的に形成する構造[7]に類似した構造があり、それが、コガネムシの表皮の光学特性に影響を与えているという。幸いに、論文で取り上げられているのと同種の虫が手元にいたので、早速観察することにした。虫はプラチナコガネのgloriasaである

これは、大昔に志賀昆虫普及社で買った物。随分と足がもげてしまい、なかなか悲し状況だけれど、コレクションのほぼ1号なので、思いで深い虫である。それが、こんなところで活躍するとは思わなかったというべきか、それとも、手元にありながら顕微鏡で覗いたことのない迂闊さを嘆くべきか…。それはともかく、それを顕微鏡で覗くと

と六角形を基本とする構造に埋め尽くされている。ちなみに、これはgoriasaの緑色の部分で、金色の筋の部分にはこのような構造はなく、均一に見える。この構造は光学的に単純ではない。照明の見込み角をかえたり、照明範囲を変えたりすると、思いがけない変化を示す。まず、照明範囲を変えてみよう
真中の明るい部分のみが落射で照明されている領域である。驚くべきことに、照明されている範囲外からの戻り光がある。その様子を眺めるとセルの片側にあたった光はセルの反対側から戻ってきている様である。もちろん、片側のみ光が当たっているセルの光が当たっている側からの戻り光もあるが、色が鮮やかでない。それに対して反対側からの光の色はより鮮やかである。これは、反射に複数の機構があることを示唆している。そして、コレステリックの特性反射は特定の波長しか戻さないことを考えると、光が当たっていない側からの戻り光がコレステリック反射由来であることを示唆している。続いて、照明の見込み角を変えた写真である。
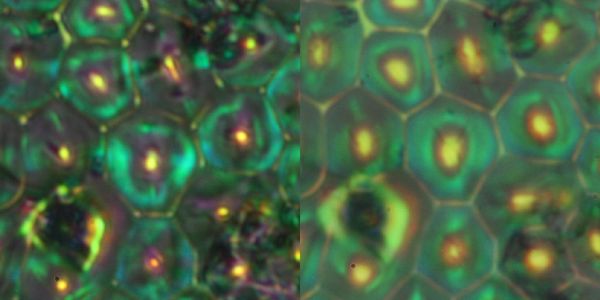
左が低NA、右が高NAである。低NAだと戻り高がセルの周辺部に限られる。これは、セルの周辺の方がコレステリックの層構造の法線方向が、より表面に垂直になっていることを示唆している。続いて、光を落射照明ではなく、斜方から入射したもの。所謂暗視野照明に相当しており、表面が平なら光はまったく対物レンズに入らないはずである。
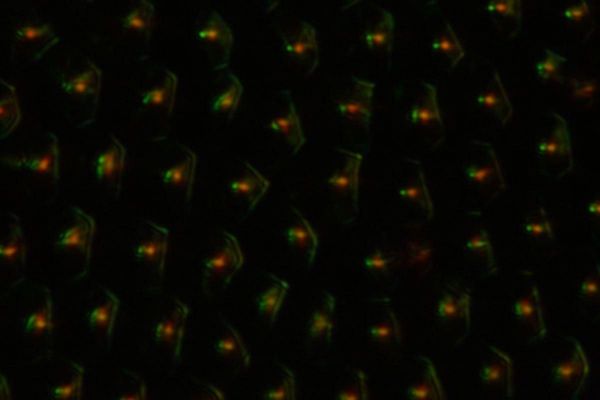
ところが、写真で明らかなように、入射光の一部は対物レンズに入射している。参照文献6で示されているような表面のフォーカルコニック構造があれば、これらの光学特性は多分説明がつくとは思う。ただし、その成因が自発的な自由界面での液晶の構造形成かと言われると、他のコガネムシもついでに顕微鏡で覗いた感じからは、肯定しきれないでいる。
ただ、それでも、このような構造をとる生物学的なメリットは容易に判る。このような構造があると、光線状態によらず見え方が一定するのだ。次に示すのはgloriasaとオプティマを内側を黒く塗った箱にいれて、カメラ側からストロボ一灯で撮影したものである。gloriasaは色を見せているのに対して、オプティマは白銀色を失い鞘翅の内側が見えてしまっている。オプティマの白銀色は周囲の光の映りこみなので、映りこみがないとこのようになってしまう。昆虫写真家の海野和男さんが小諸日記の中で自然の中で金属光沢のコガネムシをフラッシュ撮影すると色を失うと記しているけれど[8]、これはまさにこの事情によるもので、フラッシュ光は局所的でオプティマの鞘翅に映しこまれるはずの周囲までは照らしてくれないので金属光沢を失ってしまうのである。

コガネムシ上科におけるコレステリックタイプの分布
上に示したように、いろいろな虫がコレステリックタイプの反射を示す。しかし、誤解しないで頂きたいのは、これらの虫は円偏光板を持って虫屋さんに行って選んで買ってきたものであり色の有る虫をかってきても、多くはコレステリックタイプではない。しかし、経験的に、虫屋さんでも、どの箱を探せばコレステリックタイプの虫に出会えるかは分かってきている。それは、基本的にはコガネムシのたぐいである。実は、2009年11月の構造色研究会でコガネムシに関する話題を提供したときに、参加者の方からコガネムシ以外の情報を伺ったのだけれど、それについては、きちんとした報告が誰かの手によってなされるまでは、ここでは出さないつもりである。コガネムシ以外としては、世の中には玉虫の反射も円偏光とする本もあるのだけれども、東南アジア産と国産の玉虫で確認した範囲でコレステリック型ではなかった。ちなみに、本に書いた本人も……………現在は深くか浅くかは知らないけれど反省しているようである。なお、2008年に、海棲生物の蝦蛄の一種が円偏光を感知できる目を持っていることが明らかにされているが[9]、その蝦蛄の発色が円偏光かは確認されていないように思う。
さて、コガネムシであるが、次の図に示すようにコガネムシ上科(広い分類でのコガネムシ系の甲虫)の中に中生代に分離したと考えられる中生代系統群と呼ばれる12種類の科がある[10]。これらは狭義のコガネムシではない。そして、狭義のコガネムシとは中生代に上述の12種類から分離した1群の虫たちで、その下に12種類の亜科(科よりは狭い分類)がある。科や亜科の下にはさらに属がありその下に個々の種がつながっている(らしい。このあたりの生物学の分類方法に関しては専門外で、また、真面目に調べていない)。
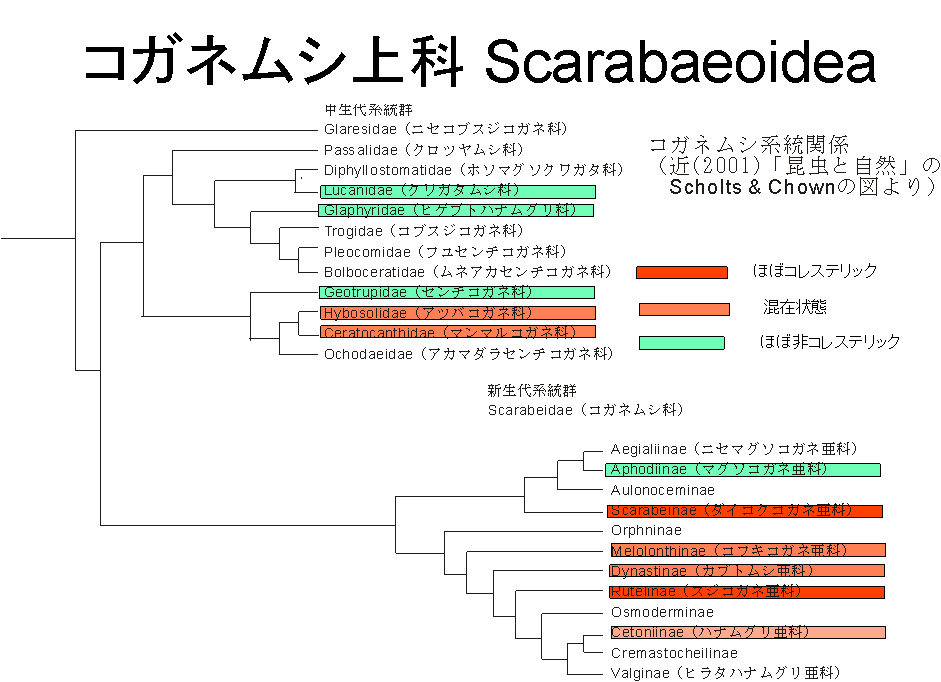
図では、系統樹の中にコレステリックタイプが見つかっているものや存在していないものを書き込んでいるけれども、これは、「コガネムシ研究会」の方々にご協力いただいて調べたもの(と自分で確認したものを)まとめたものである。もとの結果を以下に紹介しておく。
コガネムシ上科コレステリック、非コレステリックタイプ調査結果(2004年1月25日版)(この結果は、コガネムシ研究会の、佐野様、越智様、近様からのご報告と藤岡様のコレクション(の一部を)見せて頂いてまとめたものです。ここに感謝の意を表します。有り難うございました。 なお、表記等にミスもあるかと思いますが、その責は石川にございます。誤りがありましたら、石川までお教え頂ければ幸いです。また、分類はSholtsらの分類に従っております。)。なお、コレステリックタイプのコガネムシは今まで観察されているものは、全て同じ円偏光を反射しており、逆のものは見つかっていない。
| 科(family) | 亜科(subfamily) | 連(tribe) | 属(Genus) | 属和名 | コレステリックタイプ | 非コレステリックタイプ |
| Glaresidae(ニセコブスジコガネ科) | ||||||
| Passalidae(クロツヤムシ科) | ||||||
| Diphyllostomatidae(ホソマグソクワガタ科) | ||||||
| Lucanidae(クワガタムシ科) | ||||||
| Glaphyridae(ヒゲブトハナムグリ科) | ヒゲブトハナムグリは下に示した種の他、何匹が見せて頂いたが、何れも非コレステリックタイプであった。 | |||||
| Glaphyridae(ヒゲブトハナムグリ科) | Amphicoma | splendens(オオヒゲブトハナムグリ) | ||||
| trogidae(コブスジコガネ科) | ||||||
| Pleocomidae(フユセンチコガネ科) | ||||||
| Bolboceratidae(ムネアカセンチコガネ科) | ||||||
| Geotrupidae(センチコガネ科) | センチコガネは下記に示すように、観察された限りでは非コレステリックタイプである。 | |||||
| Geotrupidae(センチコガネ科) | Anoplotrupes | stercorosus(ブルー) | ||||
| Geotrupidae(センチコガネ科) | Ceratophyus | gopherinus(薄い紫) | ||||
| Geotrupidae(センチコガネ科) | Enoplotrupes | sharpi(紫) | ||||
| Geotrupidae(センチコガネ科) | Geotrupes(Onychotrupes) | semiocacas(金) | ||||
| Geotrupidae(センチコガネ科) | Lethrus | karelini(銅色) | ||||
| Geotrupidae(センチコガネ科) | Mycotrupes | gaigei(銅色) | ||||
| Geotrupidae(センチコガネ科) | Phelotrupes | auratus(オオセンチコガネ:紫・ミドリ・プルー) | ||||
| Geotrupidae(センチコガネ科) | trypocopris | vernalis(薄い紫) | ||||
| Hybosolidae(アツバコガネ科) | ||||||
| Hybosolidae(アツバコガネ科) | Phaeochrous | philippinensis(前胸背に弱い紫銅光沢),emarginatus(弱い紫銅光沢) | ||||
| Ceratocanthidae(マンマルコガネ科) | ||||||
| Ceratocanthidae(マンマルコガネ科) | Ceratocanthus | volvox(銅) | ||||
| Ceratocanthidae(マンマルコガネ科) | Eusphaeropeltis | aurora(グリーン) | ||||
| Ochodaeidae(アカマダラセンチコガネ科) | ||||||
| Scarabaeidae(コガネムシ科) | ||||||
| Aegialiinae(ニセマグソコガネ亜科) | ||||||
| Aphodiinae(マグソコガネ亜科) | ||||||
| Aphodiinae(マグソコガネ亜科) | Aphodius | contaminas(薄い銅) | ||||
| Aulonocneminae | ||||||
| Scaraabaeinae(ダイコクコガネ亜科) | ダイコクコガネは下に示すように、コレステリックタイプである。結構いろいろな色があるけれども、それらが全てコレステリック発色で出ている。ニジイロダイコクコガネのようにカラフルなものもいる。 | |||||
| Scaraabaeinae(ダイコクコガネ亜科) | Dichotomiini | Bolbites | onitioides(赤、グリーン) | |||
| Scaraabaeinae(ダイコクコガネ亜科) | Caccobius | fortus(薄い銅) | ||||
| Scaraabaeinae(ダイコクコガネ亜科) | Canthonini | Canthon | vividis(グリーン) | |||
| Scaraabaeinae(ダイコクコガネ亜科) | Canthonini | Cassolus | peninshularis(薄いグリーン) | |||
| Scaraabaeinae(ダイコクコガネ亜科) | Dichotomiini | Chalcocopris | hespera(オリーブ) | |||
| Scaraabaeinae(ダイコクコガネ亜科) | Onitini | Chironitis | hungaricus(グリーン) | |||
| Scaraabaeinae(ダイコクコガネ亜科) | Canthonini | Deltochilum | gibbosum(薄い銅) | |||
| Scaraabaeinae(ダイコクコガネ亜科) | Dichotomiini | Gromphas | aeruginosa(グリーン) | |||
| Scaraabaeinae(ダイコクコガネ亜科) | Gymnopeulini | Gymnopleurus | aciculatus(ブルー), coerulescens(紫), | |||
| Scaraabaeinae(ダイコクコガネ亜科) | Canthonini | Hansreria | affinis(グリーン) | |||
| Scaraabaeinae(ダイコクコガネ亜科) | Scarabaeini | Kheper | festivus(グリーン), sanctus(グリーン), purpurascens(紫) | |||
| Scaraabaeinae(ダイコクコガネ亜科) | Oniticellini | Liatongus | militaris(グリーン) | |||
| Scaraabaeinae(ダイコクコガネ亜科) | Phanaeini | Megaphanaeus | ensifer(グリーン) | |||
| Scaraabaeinae(ダイコクコガネ亜科) | Canthonini | Megathopa | bicolor(赤銅) | |||
| Scaraabaeinae(ダイコクコガネ亜科) | Onitini | Onitis | humerasus(グリーン) | |||
| Scaraabaeinae(ダイコクコガネ亜科) | Onthophagus | watanabei(紫銅) | ||||
| Scaraabaeinae(ダイコクコガネ亜科) | Pachylomera | conspicillatum(グリーン) | ||||
| Scaraabaeinae(ダイコクコガネ亜科) | Oniticellini | Paroniticellus | festiuus | |||
| Scaraabaeinae(ダイコクコガネ亜科) | Phanaeini | Phanaeus | damon(グリーン) | |||
| Scaraabaeinae(ダイコクコガネ亜科) | Scarabaeini | Scarabaeus | radama(薄い銅色) | |||
| Scaraabaeinae(ダイコクコガネ亜科) | Sysiphini | Sisyphus | thoracicus(薄いグリーン) | |||
| Orphninae | ||||||
| Melolonthinae(コフキコガネ亜科) | 光沢や色を示す虫がすくないが、ビロードコガネともう一種(?アボゴニア)はコレステリックタイプであった。分類的にダイコクコガネとカブトムシの間なのでコレステリックタイプであるのはめでたいことである。 | |||||
| Melolonthinae(コフキコガネ亜科) | ||||||
| Dynastinae(カブトムシ亜科) | カブトムシは、色がなく光沢もない虫がほとんどなので、コレステリックタイプか非コレステリックタイプかは謎だった。系統樹的にはコレステリックタイプになって欲しいところであったが、某昆虫屋さんで下記のbicuspisを見つけてコレステリックタイプであることが確認した時は嬉しかった。残念ながら高い虫でコレクションにつれて帰れなかったけれど。 | |||||
Dynastinae(カブトムシ亜科) |
Aegopsis | bicuspis(鈍い緑青) | ||||
| Rutelinae(スジコガネ(キンコガネ)亜科) | スジコガネはほとんどコレステリックタイプである………。が、クワガタコガネ(東南アジア)と南米のスジコガネ(plusiotisを含む?)の何種類かが一見非コレステリックタイプである。ただし、これらのスジコガネは、非コレステリックタイプではなくresplendensのように両方の円偏光を反射している可能性がある(下の方の写真とコメント参照)。まあ、基本的にはコレステリックタイプと考えて良い。 | |||||
| Rutelinae(スジコガネ(キンコガネ)亜科) | Anomala | albopilosa(アオドウガネ), cupera(ドウガネブイブイ、鈍銅) | ||||
| Rutelinae(スジコガネ(キンコガネ)亜科) | Mimela | confusius(イシガキコガネ) | ||||
| Rutelinae(スジコガネ(キンコガネ)亜科) | Plusiotis | optima(白銀),batesi(薄緑銀), resplendens(真鍮色、両円偏光を反射) | ||||
| Osmodeminae | ||||||
| Cetoniinae(ハナムグリ亜科) | ハナムグリ亜科は混沌としている。亜科の中でもバラバラであるどころか、同じ属の中でもコレステリックタイプと非コレステリックタイプが存在したりする。例えばCallistethusに属すると思われている南方系の虫はコレステリックタイプで北方系のものは非コレステリックタイプであった。 大きなくくりとしてはカナブン系は非コレステリックタイプであることが多いのだが、例外も少なくなく、今後の検討が必要だろうと思っている。 | |||||
| Cetoniinae(ハナムグリ亜科) | Coptomia | マラガシーハナムグリ属 | laevis | |||
| Cetoniinae(ハナムグリ亜科) | Cotinis | ヒシムネハナムグリ属 | subviolacea | |||
| Cetoniinae(ハナムグリ亜科) | Cetonia | ハナムグリ属 | aurata | |||
| Cetoniinae(ハナムグリ亜科) | Protaetia | ハナムグリ属 | montana,orientalis,affinis | |||
| Cetoniinae(ハナムグリ亜科) | Chiloloba | ハナムグリ属 | acuta | |||
| Cetoniinae(ハナムグリ亜科) | Stenopisthes | ゴウシュウハナムグリ属 | frenchi | |||
| Cetoniinae(ハナムグリ亜科) | Lomaptera | ゴウシュウハナムグリ属 | linae,burgeoni | |||
| Cetoniinae(ハナムグリ亜科) | Ischiopsopha | ゴウシュウハナムグリ属 | jamesi,ritsemae | |||
| Cetoniinae(ハナムグリ亜科) | Hemipharis | ゴウシュウハナムグリ属 | insularis | |||
| Cetoniinae(ハナムグリ亜科) | Anacamptorrhina | ゴウシュウハナムグリ属 | ignipes | |||
| Cetoniinae(ハナムグリ亜科) | Theodsia | カブトハナムグリ属 | howittii,viridiaurata | |||
| Cetoniinae(ハナムグリ亜科) | trigonophorus | カナブン属 | riaulti | feae | ||
| Cetoniinae(ハナムグリ亜科) | Rhomborrhina | カナブン属 | fuscipes,gigantea | |||
| Cetoniinae(ハナムグリ亜科) | Raceloma | カナブン属 | natalensis | |||
| Cetoniinae(ハナムグリ亜科) | Ptychodesthes | カナブン属 | gratiosa | |||
| Cetoniinae(ハナムグリ亜科) | Plaesiorrhina | カナブン属 | watkinsiana | rufonasuta | ||
| Cetoniinae(ハナムグリ亜科) | Neoscelis | カナブン属 | dohrni | |||
| Cetoniinae(ハナムグリ亜科) | Narycius | カナブン属 | opalus | |||
| Cetoniinae(ハナムグリ亜科) | Mystroceros | カナブン属 | macleayi | |||
| Cetoniinae(ハナムグリ亜科) | Jumnos | カナブン属 | rucheri | |||
| Cetoniinae(ハナムグリ亜科) | Ingrisma | カナブン属 | burmanica | |||
| Cetoniinae(ハナムグリ亜科) | Heterorrhina | カナブン属 | macleayi,elegans | |||
| Cetoniinae(ハナムグリ亜科) | Dymusia | カナブン属 | variabilis | |||
| Cetoniinae(ハナムグリ亜科) | Dicronorhina | カナブン属 | micans | |||
| Cetoniinae(ハナムグリ亜科) | Cyphonocephalus | カナブン属 | olivaceus | |||
| Cetoniinae(ハナムグリ亜科) | Anomalocera | カナブン属 | subopaca | |||
| Cetoniinae(ハナムグリ亜科) | Paraplectrone | エグリハナムグリ属 | crassa | |||
| Cetoniinae(ハナムグリ亜科) | Glyptothea | エグリハナムグリ属 | whiteheadi | |||
| Cetoniinae(ハナムグリ亜科) | Chaloteomima | エグリハナムグリ属 | leavis | |||
| Cetoniinae(ハナムグリ亜科) | Chalocohea | エグリハナムグリ属 | neglecta | |||
| Cetoniinae(ハナムグリ亜科) | Callistethus | 南方系2種(緑) | 北方系1種(緑) | |||
| Cremastocheilinae | ||||||
| Valginae(ヒラタハナムグリ亜科) | ||||||
| trichiinae | ||||||
| Euchirinae(テナガコガネ亜科) | テナガコガネは観察した範囲ではコレステリックタイプである。飛翔羽の上の鞘の部分は黒色でコレステリック反射がみられない場合も、その場合でも頭部の甲がコレステリックタイプの反射を示していることがある。 | |||||
| Euchirinae |
進化系統とコレステリック・非コレステリックの関係を見ようと思い立ったのは、物理化学的なイメージからすると、コレステリックタイプと誘電体多層膜タイプでは大きな違いがあるからである。コレステリックタイプは基本的に1種類の物質からできている。さなぎの時点で、どのようにラセン構造が形成されるのかは分からないが、通常のコレステリック液晶の場合は、適当な温度あるいは溶液系の場合は濃度で勝手にラセン構造が形成される。何もないとラセン軸の方向はランダムなのだけれど、適度な界面があれば配向して揃ったラセンを作ることは比較的容易である。一方の誘電体多層膜タイプは基本的に2種類の物質からできている。2種類の混合系が周期構造を作る物理化学的な機構としてはスピノーダル分解が知られているのだが、スピノーダル分解では3次元的な構造となってしまう。とても、積層多層膜ができるとは思いがたい。誘電体多層膜タイプの場合には、物理化学的な機構以外の生物学的な機構が構造形成に関与しているのではないかと思える。つまり、2つのタイプで人間にとって見え方は似ていても、機構的には大きく異なっているのではないのかと思えるのだ。こうしてみると、進化的にも、誘電体多層タイプとコレステリックタイプの間にハッキリとした境界があっても良さそうだと思ったのである。
残念ながら、系統樹との関連に関しては大雑把な規則性はあるものの、例外も多く、またScholtsらによる系統関係とは必ずしも対応しておらずに、困っている。
もっとも、最近のミトコンドリアDNAによる系統樹の研究(九州大学荒谷氏らによる)によると、これまでスジコガネやカブトムシと類縁と考えられていたクワガタコガネは、むしろハナムグリに近いという結果が得られているらしい。これは、コレステリック・非コレステリック分類から言えばきわめて有り難い結果である。なにしろ、スジコガネもカブトムシもコレステリックタイプであるので、その中で非コレステリックタイプの一群がいるのは、頭痛の種だったからである。しかし、ハナムグリの近縁となれば、クワガタコガネの一族揃って非液晶タイプであることには問題はない。そして、反射機構の違いがDNA分類を支持するという意味でも、互いにめでたい結果であろう。
また、上記の結果をみていると、ハナムグリの中での属間の類似度が他の亜科に比べて低い印象がある。このことは、実はハナムグリ亜科は分類学的には亜科より上のランクであることを示唆しているように個人的には感じられている。そしてまた、Scholtsの分類と比べると、より上位で分科を起こした一族となっており、このこともハナムグリ一族の中にコレステリックタイプと非コレステリックタイプが混在する結果を説明するのに好都合である(ある形態を獲得したあとに、時間が経つほど、その形態は安定になるだろうと勝手に信じている。それ故、Scholtsの分類のように、ハナムグリの分岐が、ダイコクコガネ(コレステリック)との分岐、そしてスジコガネ(コレステリック)との分岐の後に起こっているとしたら、コレステリック形態は十分に安定しているはずで、亜科の中でバラバラになる理由が存在するとは思えないところであった。しかし、ハナムグリの分岐が、古い物なら(とはいえ、ダイコクコガネとの分岐よりは後のようだけれど)、形態の安定は不充分でバラバラになることも、あってもいいかもしれない。
何故コガネムシは色を示すのか
コガネムシが構造色により鮮やかに光り輝く生物学的な意味については、未だに解明されていないような印象を受ける。「昆虫と自然」に掲載された故知の記事によると構造色の生物学的な役割について、
- 体温調整説
- 性選択説
- 同性内選択
- 異性間選択
- 捕食者適用仮説
- 補食対象の位置確認の困難化
- 隠蔽色
- 警告色
- コントラスト変化説
などがあるとされている。故知は上記のうち、体温調整説と警告色仮説に関して簡単な実験を行い、否定的な結果を得ている[11]。
体温調整説というのは、直射日光下で太陽光により昆虫の体温が上がりすぎるのを防ぐために構造色により太陽光を反射しているという説である。確かに構造色では光の吸収なく反射が起こる。しかしコレステリックタイプである限り、ある波長領域の50%の光を反射するのが限界である。さらに黒色色素を持っているということは反射できなかった光は有効に吸収している分けで、体温上昇を防ぐこととは昆虫の構造が矛盾している。ただし、金属光沢を有するプラチナコガネ、特にレスプレンデンスに関しては、太陽光を可視の広い範囲に渡って、50%以上の反射率を有しており、また羽の下層の黒色色素を持っていないので体温調整説に適合する構造となっている。しかし、レスプレンデンスにおいても、体温調整説は妥当ではないだろう。金属光沢のプラチナコガネ類は南米原産であるがT.イッセイ氏のエクアドルにおけるプラチナコガネ採集記によれば、生息地の温度は20摂氏程度、湿度は80%の森林で飛来時間は19時から21時の夜間である[12]。太陽光による温度上昇が問題となるような生息条件ではないようなのであるから(その後、藤岡さんに伺ったところでは、プラチナコガネの生息域は基本的には高温多湿の密林で昼間も活動するとのことであった。ただし、生息域からして太陽光を森林等で遮蔽することは容易であり、やはり体温調整説は生息域の環境からは否定できるとのことであった。)
性選択説が成立するためには、コガネムシが円偏光を認識できることが必要条件となるが、これは、現時点では確認されていない。また、左右両円偏光を反射するコガネムシでは、この仮説は成立しない。
警告色仮説とは、毒のある生物がそれ故に他の動物にはっきりと目につくような色彩を取ることがあるというもので、これにより他の生物に対して警告をしているとするものである。コレステリックタイプの虫に関して、警告色仮説が成立するかというと故知の実験の範囲ではコレステリックタイプのコガネムシも毒を有している気配はなく、また鳥類にも隔てなく食されるようで、一般的には当てはまらないようである。
上記以外で保護色(隠蔽色)である可能性については緑色系のコガネムシについては、漠然とした印象としてはある程度は当てはまるように思える。保護色に関して、少しばかり怪しげな議論を進めてみよう。
コレステリックタイプと誘電体多層膜タイプを比べると、コレステリックタイプでは反射帯でも反射率の最大値が50%であるのに対して、誘電体多層タイプでは反射率が100%となりうることである。つまり、平均してコレステリックタイプの方が平均して暗くなっている可能性がある。これが、実際の植物の緑の反射率と比べた場合にどちらが有利であるかは…微妙なところであるかもしれないが。
ところで、植物の中には深い緑の他に新緑のような明るい浅い緑色のものもある。では、コレステリックタイプではこの緑が出ないかというと、そうでもない。羽の下層に黒色の色素層があると確かに反射率の最大は50%で深い緑になる。しかし、下層に色素層がなく、羽を通過した光が下層で反射して戻ってくる場合には全体に明るくなって色も浅くなる。実際に50%程度の緑と、ベースが乗った淡い緑を作ったものを下に示す。

実際、コガネムシの中には淡い緑で偏光フィルターで見ると色素層が欠落しているように見えるものがある。自然界はまったく器用なものである。
では、金属光沢のコガネムシはどうかというと、その金属光沢故に回りの景色にとけ込む可能性がある。

写真は、コガネムシの代わりに直径20mmほどの真鍮の球体を草の根本において写真撮影したものである。空が3カ所写り込んでいるので球の存在が分かるが、それ以外の部分は回りの風景が写り込んでいて風景の中にとけ込んでいるような気がするではないか。実は南米の熱帯雨林には金色に輝く光沢の繭を作る蝶がいる。身動きがとれずに木にへばりついている状態で、そんなに金属光沢で大丈夫なのかと不思議なのだが、熱帯雨林の中では風景にとけ込んで、時には木漏れ日のようでもあり保護色になっているらしい。金属光沢のコガネムシも同様な意味で保護色になっている可能性がある。
付録:円偏光と円偏光板
積層タイプとオパールタイプの機構の説明は、そのうちに気力と余力ができたらとして、ここでは、コレステリックタイプの特徴と機構の説明をすることにしよう。このタイプの反射光の特徴は円偏光となっていることである。ここで円偏光という少しばかり耳慣れない言葉が出てきたので先に進む前にその説明をしておこう。光は電磁波であり横波の一種である。この横波の振動方向が揃った状態を偏光という。
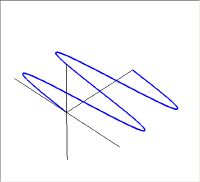
図に振動方向が水平(青)と垂直(赤)方向にある偏光を示した。この偏光は振動面を進行方向から見たときに直線上を振動することになるので直線偏光と呼ばれている。次ぎにこの2つの直線偏光の重ね合わせを考えてみよう。
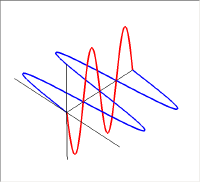
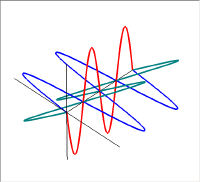
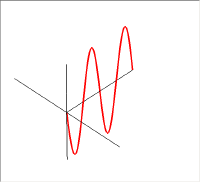
左の図が上の2つの直線偏光を重ねて描いた物で、右の図は進行方向の軸の同じ位置での振幅をベクトル的に足し合わせたものである。新たにできた緑色の振動も直線偏光で、その振動方向は垂直・水平軸から45度傾いている。この絵では足し合わせる2つの波の位相が同じであった。しかし、2つの波の位相が異なると足し合わせた結果はもはや直線にはならない。

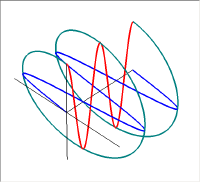
この図では赤い波の位相を青い波に対して90度違えてある。この2つの波を足し合わせると右側の図の緑の線のように振動は螺旋状になる。このラセンを進行方向からみると円となるので、このような状態を円偏光と呼ぶ。
円偏光には右円偏光と左円偏光がある。右円偏光と左円偏光ではらせんの掌性(キラリティ)が異なっている。

足し合わせる2つの波の位相が90度以外の場合(や位相差は90度だけれど振幅が異なっている場合)は、振動の軌跡は進行方向からみて楕円となるので楕円偏光と呼ばれている。直線偏光と円偏光は楕円偏光の特別な場合である。
太陽光、電球や蛍光灯からの光は一般的には特別な偏光状態とはなっておらず、このような状態の光を自然光と呼ぶ。甲虫の羽で光が反射する場合に、誘電体多層膜タイプやオパールタイプでは入射方向によっては直線偏光成分が発生することはあるけれども、円偏光成分が出てくることはない。ところが、コレステリックタイプの場合にはきわめて例外的な場合を除き円偏光となる。
偏光を作り出すのに、かつては複屈折性のある方解石を用いたプリズムなどが使われていた。その後、ランド博士(インスタント写真の発明者としても有名)によりフィルム偏光板が発明され、現在では多くの用途にフィルム偏光板が使われている。直線偏光板は方向の揃った沃素の針状微結晶を含んだ高分子フィルムで、針状結晶の方向に平行な電場成分の光を選択的に吸収して、自然光の中から片方の偏光の光のみを取り出している。通常の直線偏光板は可視領域で性能を発揮するように作られており、紫外光や赤外光は透過しなかったり偏光度が低下したりする。直線偏光板の主な民生用途は釣り用のサングラスやカラー写真用のコントラスト向上フィルターなどであったが、最近では液晶表示装置に数多く用いられている。なお、直線偏光板には裏表はない。 円偏光板は直線偏光板に複屈折性のあるプラスチック位相差フィルムを(偏光子の光軸とフィルムの光軸を45度ずらして)貼り付けた積層構造フィルムである。自然光は円偏光板の直線偏光部分を透過して、一旦直線偏光になった後、位相差フィルムを通過して円偏光になる。右円偏光板と左円偏光板では、直線偏光子の軸に対する位相差フィルムの軸の関係が逆になっている。
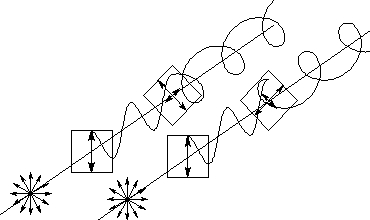
図:円偏光板の構造 円偏光板は直線偏光板(手前の矢印が1本の板)と1/4波長板(奥の直交した矢印の板)を張り合わせたものである。ここでは、分かりやすいように2枚の板を離して描いている。 無偏光の光は直線偏光板を通って直線偏光になる。その後1/4波長板を通って円偏光になる。右円偏光になるか、左円偏光になるかは、1/4波長板の軸が直線偏光板の軸の対してどちらに45度傾いているかで定まる。
円偏光板は直線偏光板と位相差板を積層した構造になっているので裏表がある。直線偏光側から自然光を入れると円偏光が出てくるが、位相差板側から自然光を入れても、直線偏光しか出てこない。これは自然光が位相差フィルムを通過しても、それぞれに位相が変化するだけで全体としては自然光のままであるために(つまり、特別な規則性が出現しない。位相差フィルムは、入射光に特別な規則性がある場合にのみ意味のある働きをする。)、引き続き通過する直線偏光板で直線偏光に変換されるのみだからである。
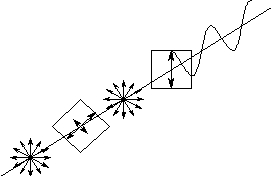 図:円偏光板を逆向きで使った場合
1/4波長板の側から無偏光を入れると、1/4波長板を通っても(位相が変調を受けるだけで)無偏光のままである。その光が直線偏光板を通って直線偏光となって出てくる。つまり、円偏光板を単独で逆向きで使うと直線偏光板のような動きをする。
図:円偏光板を逆向きで使った場合
1/4波長板の側から無偏光を入れると、1/4波長板を通っても(位相が変調を受けるだけで)無偏光のままである。その光が直線偏光板を通って直線偏光となって出てくる。つまり、円偏光板を単独で逆向きで使うと直線偏光板のような動きをする。
それに対して位相差フィルム側から円偏光を円偏光板に入射すると、位相差フィルムにより円偏光は直線偏光に変換され、その直線偏光の軸は円偏光板の直線偏光子の軸と一致するか直交するかとなり、光は透過又は遮断される。具体的には位相差フィルム側から左円偏光板に左円偏光を入れると光は透過し、右辺偏光を入れると光は遮断される。
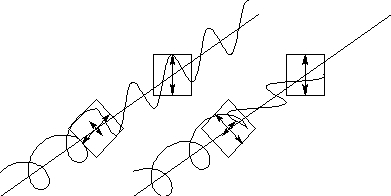 図:円偏光板への円偏光の入射
左と右で円偏光板の符号が異なっている。左では入射した円偏光はそのまま通過するが、右では直線偏光板で遮断され、光は透過しない。
逆に、直線偏光板側から円偏光をいれると、円偏光が直線偏光板により直線偏光に変換され、それから位相差板を通るので、(最初の光よりも強度が半分程度以下に弱くなった)円偏光が出てくることになる。この場合は左円偏光板に左円偏光を入れても右円偏光を入れても出てくる光の強度に変化はない。
図:円偏光板への円偏光の入射
左と右で円偏光板の符号が異なっている。左では入射した円偏光はそのまま通過するが、右では直線偏光板で遮断され、光は透過しない。
逆に、直線偏光板側から円偏光をいれると、円偏光が直線偏光板により直線偏光に変換され、それから位相差板を通るので、(最初の光よりも強度が半分程度以下に弱くなった)円偏光が出てくることになる。この場合は左円偏光板に左円偏光を入れても右円偏光を入れても出てくる光の強度に変化はない。
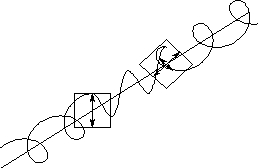
図:直線偏光板側からの円偏光の入射 直線偏光板側から円偏光を入射しても、直線偏光板で直線偏光に変換された後に1/4波長板を通って円偏光になるのみである。無偏光をいれるのと状況はまったく変わらない。
なお、円偏光板についている位相板は厳密には一つの波長でのみ直線偏光を円偏光に変換する。それ以外の波長では楕円偏光(円偏光と直線偏光が混ざったような状態)に変換する。 円偏光板を通った光は2つの直線偏光成分を持っているので、見た目には無偏光状態と同様に見える。一部の一眼レフカメラでは直線偏光の方向に敏感な光学系を有しており、このようなカメラ用の偏光フィルターは円偏光板を用いている。また、デジタルカメラも複屈折を利用した光学素子を用いており、通常の偏光板を用いると問題が生じるのでデジタルカメラ用の偏光フィルターも円偏光板となっている。円偏光板はフィルターの他に、コンピュータ画面の写り込み防止フィルターにも使われている。
人間の目は実質的に偏光を検出できないので、円偏光板をいくら眺めていても裏表の確認はできない。しかしながら、円偏光板の裏表を簡単に確認する方法がある。反射のある物の上に円偏光板を置いてみればよいのである。実は、円偏光は、ある例外を除いて反射したときに左右が入れ替わる。つまり、左円偏光は鏡などで反射すると右円偏光になり、右円偏光は左円偏光になる。このため、鏡に左円偏光板をくっつけると、もし、位相差板が鏡側にあれば、円偏光板を透過した光は左円偏光になり、鏡面で反射されて右円偏光になって戻ってくる。そして、右円偏光は左円偏光板を透過できないので、暗くなる。逆に直線偏光子が鏡側の場合には、偏光子を通過した直線偏光は鏡で反射されても、同じ直線偏光のままで、そのまま偏光子を透過して(位相差板で円偏光に変えられて)戻ってくるために、暗くはならない。なお、偏光板と反射面との距離が空きすぎていると、偏光板を透過せずにものに当たった光がやってくるので、正しい方向でもそれほど暗くならない。なるべく密着させるのが区別するコツである。
左右どちらか判らなくなった円偏光板を区別するには、その辺のコガネムシ類で緑に色付いているものを円偏光板を通して観察すればよい。どちらの偏光板でも何事も起こらなかったら、別の種類のコガネムシを見てみること。そのうち、片方の偏光板を通すと色が劇的に黒くなる虫が見付かる。色が劇的に黒くなった方が右円偏光板で、そうならない方が左円偏光板である(ただし、これは実験した範囲であり、例外的なコガネムシがいるかもしれない)。
右左のフィルム円偏光板はポラロイド社から発売されていたが、同社の倒産にともない、一方の供給が途絶え、また価格の上昇が起こった。価格は2003年当時と比較して高くなっているが、現在では美舘イメージングで比較的容易に入手出来る。
参考文献
1,Gold bugs and beyond: a review of iridescence and structural colour mechanisms in beetles, A. E. Seago, P. Brady, J-P Vigneron and T. D. Schultz, J. R. Soc. Interface, Vol. 6, (2009)S165.
2. Friedrich Reinitzer, (1888)421
3.Michelson, A. A.Phil. Mag. 21(1911) 554.
4.Cuticle reflectivity and optical activity in scarab beetles: the role of uric acid. S. Caveny , Proc. R. Soc. B V178(1971)pp.205.
5.
6.Structure Origin of Circularly Polarized Iridescence in Jweeled Beetles, V. Sharma, et al., Science 325(2009)P449.
7.R. Meister et al.,Phys. Rev. E 54, (1996)3771.
8.http://eco.goo.ne.jp/nature/unno/diary/200812/1228881127.html
9.Circular Polarization Vision in a Stomatopod Crustacean:T. H. Chiou1,S. Kleinlogel, T. Cronin1, R. Caldwell, B. Loeffler, A. Siddiqi, A. Goldizen and J. Marshall:Current Biology, V18(2008) 429-434
10.近 雅博昆虫と自然:36(10)(2001)15
11.古知 新昆虫と自然:36(10)(2001)5
12.T.イッセイ:クワタ No16(2003)95.