甲虫の構造色(コガネムシを中心に)
自然界に見られる多様な色彩は、特定の波長の光を吸収して発色しているもの(吸収色)と、光の吸収のない物理的な機構により発色しているもの(構造色)があります。
甲虫の中には様々な色彩の鞘翅を持つものがあり、それらの色彩には構造色であるものも数多くあります。
甲虫の構造色の機構をA. E. Seagoらは以下の4つに整理しています[1]。
- 誘電体多層膜
- 3次元フォトニック結晶
- 回折格子構造
- 散乱体
誘電体多層膜は2つの異なる屈折率の層が積層した構造で、多層薄膜の干渉による発色です。タマムシの発色はこの機構によるものです。

3次元フォトニック結晶は3次元周期構造による光の回折による着色で、宝石のオパールと類似した着色機構です。カタゾウムシの一種がこの機構により発色しています。

3次元構造なのは写真では白く見えている部分です。手元で見つかった写真が同じ個体のものである気がしないのですが、とりあえず見つかったものを掲載します。
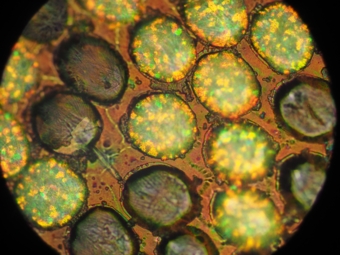
白く見える部分は丸い粒の集合体で丸い粒の中にはオパールのように着色した小片が散らばっています。
回折格子構造は鞘翅表面に周期的な溝があり、それが回折格子となり発色するもので、ある種の水生昆虫やハネカクシにみられますが、残念ながら実物を所有していないので写真を提示できません。
散乱による発色はカラフルな色ではなく、白色となりますが、まずはその例をご覧ください。

3次元フォトニック結晶を示すカタゾウムシと似た昆虫ですが、カタゾウムシではなくカタゾウカミキリです。この虫の生息域はカタゾウムシと重なっています。昆虫好きの方に教えていただいたのですが、カタゾウムシは鞘翅が融着していて飛翔できませんが、硬くて鳥が摂取しても消化できないので、鳥には襲われないのだそうです。カタゾウカミキリの方は鞘翅は融着しておらず、飛翔もできるそうですが、カタゾウムシに擬態して、鳥に食べられないようにしているのだそうです。それにしても触覚の付き方は違いますが、それ以外は類似度の高い姿をしています。そうなると、白く見える部分の構造も知りたくなるので、拡大してみてみると
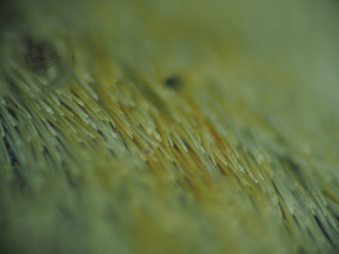
毛でおおわれていて、その乱反射で白く見えているだけで、3次元構造のかけらもないことがわかります。擬態は見た目が重要で、発色機構までは模倣する必要がないわけです。
コガネムシの発色機構
さて、A. E.
Seagoらによる区分はこの4つで終わりなのですが、彼らによって、誘電体多層タイプに分類されているコガネムシの中には、通常の誘電体多層膜とは異なった構造と光学特性のグループが存在します。

写真に示すのはコガネムシ上科の2頭の虫です。左の虫の方が少し暗めの緑ですが、まあ、外形的にも似た印象の虫です。この写真は偏光フィルターなしに撮影したものですが、右円偏光フィルターを通してみると次のようになります。

右の虫は変化がありませんが、左の虫は色味を失って暗くなっています。続いて、左円偏光フィルターを通してみてみましょう。

右の虫は変化がありませんが、左の虫は、今度は相対的に明るくなっています。右の虫は通常の誘電体多層構造による発色ですが、左の虫の発色機構は、それとは、何かが異なっているのです。
右円偏光とか左円偏光という言葉になじみのない方もいらっしゃるかと思いますので、ここで、どんなイメージのものかだけを簡単に紹介します。
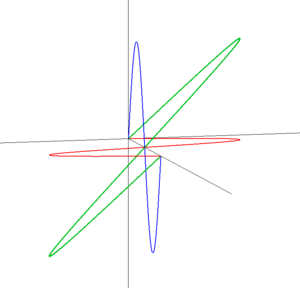 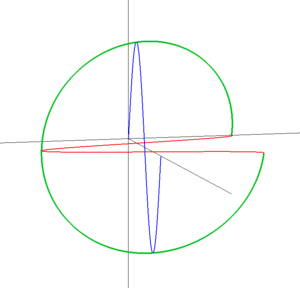
図:直線偏光(左)と円偏光(右)
左の図は斜め45度方向に振動する直線偏光です。直線偏光は2つの直交する偏光(赤と青)の重ね合わせで作り出せます。直線偏光では青い波と赤い波は位相が同じです。ここで、赤い波の位相を1/4波長だけずらした状況を考えてみましょう(右)。赤い波と青い波を重ね合わせた波は、円を描くような軌跡をたどっています。これが円偏光です。図では緑の線は右ネジの巻きになっていますが(右円偏光)、巻きが逆だと左円偏光になります。
直線偏光では特定の方向にのみ波面があるので、もう1枚の偏光子を、その波面と垂直な方向の光のみを透過するように重ねると、光が遮断されます。一方、円偏光では波面はすべての方向に回転しながら変化しているので、普通の偏光子をいれても、角度依存性はなく、偏光していない光と区別がつきません。これに対して右円偏光板は右円偏光は透過しますが、左円偏光は遮断します。そして左円偏光板は左円偏光を透過し、右円偏光を遮断します。
円偏光板を通してみた虫の写真に戻ると、右円偏光板をいれると暗くなり、左円偏光板では明るくなったということは、左側の虫からの反射光は左円偏光になっていることになります。
反射光が左円偏光になっていることはわかりましたが、どのような入射光が反射されて左円偏光になっているかは、反射光を見ただけではわかりません。円偏光の左右は鏡で反転してしまうからです。

図鏡の上に偏光板
写真は偏光板を鏡の上にのせたものを上から撮影したものです。右側のCの文字が貼ってある偏光板は普通の直線偏光板です。偏光板を通過した直線偏光が、鏡でそのまま反射されて、直線偏光板を透過してきます。左側のYの文字がはってあるものは右円偏光版です。こちらは、光が透過していません。右円偏光板を透過した右円偏光は鏡で反射すると左円偏光になってしまいます(右手を鏡に映すと左手になってしまいますよね。)。このため、右円偏光板を通れず暗くなってしまったのです。
では、先ほどの虫はどうなるのでしょう。

図:右円偏光板を直接上に置いた状態
右円偏光板を直接上に置いてみました。左側の虫は上の写真と同様に暗くなっています。そして右側の虫も暗くなりました。これは、鏡と同じように、右円偏光板を透過した光が反射する時に左円偏光になってしまったためです。続いて、左円偏光板を上において観察してみましょう。

図:左円偏光板を直接上においた状態。
右側の虫は暗いままです。これは、左円偏光板を透過した左円偏光が虫表面で反射して右円偏光になってしまったためです。一方、左側の虫は明るく色が見えています。ということは、左円偏光板を透過した左円偏光が、反射しても左円偏光のままで、左円偏光板を透過してきたことになります。
コガネムシの一族の中には、左円偏光のみを反射して、しかも反射光が左円偏光のままになるという光学特性の昆虫が存在しているのです。もちろん、目視では、左の虫が、そんな奇妙な光学特性をしていることはわかりません。それにも関わらず、1900年代初頭には、コガネムシの中に、こんな変わった光学特性を持つものがいることに気が付いた人がいて、さらに、この特性がコレステリック液晶(キラルネマチック液晶)に類似していることも認識されていました。

図:キラルネマチック液晶
図にキラルネマチック液晶の様子を示します。ネマチック液晶と同様に棒状の分子ならなる液晶で、不斉炭素を持つ分子が含まれています。そのため、分子長軸方向は空間的に均一ではなく、ある軸に垂直な面内方向の分子長軸の方向軸方向に進むにつれて、が一方向に回転しています。回転の様子を見やすくするために、層構造があるかのような図も使われることがありますが、層構造はなく、分子の重心はランダムに運動しています。分子の長軸が360度回転する周期をd、液晶の平均屈折率をnとすると、波長λがndを中心とした反射が生じます。そして反射するのはコレステリック液晶のラセンが右巻きらせんなら右円偏光のみ、左巻きなら左円偏光のみを反射し、反対の円偏光は透過します。この反射機構については、別の頁で説明します。
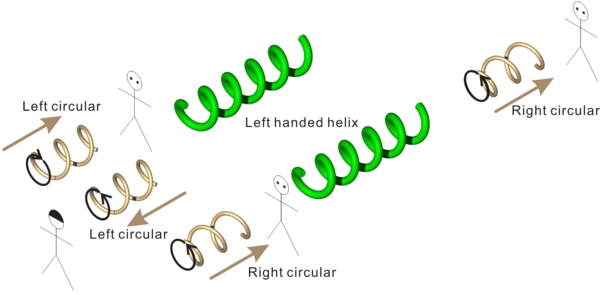
図:螺線構造と反射する円偏光
コガネムシの鞘翅は液晶状態ではありませんが、コレステリック液晶のように分子(高分子)の長手方向が螺線を描くような構造をしています。螺線軸はもちろん鞘翅に垂直な方向です。左円偏光のみを反射することから、螺線のねじれは左巻きで、有機分子の屈折率が1.5程度とすると、500nmの緑が反射する場合には、330nm程度の周期構造になっているはずです。鞘翅の断面を電子顕微鏡で観察すると、160nm程度周期の縞構造が見られます。コレステリック液晶は非極性なので、180度軸が回転した状態は0度の状態と物理的に同じです。電子顕微鏡で観察されている周期は180度回転に対応したものです。
写真に示したものはトルコのコガネムシです。鞘翅は緑ですが、胸の部分は赤色です。両者で螺線周期が異なっているか、平均屈折率が異なっているか両方とも異なっているかになっているはずです。

どのようなコガネムシが円偏光を反射するのかなどについては、昔にしるしたものがありますので、興味のあるかたは、ご覧ください。
プラチナコガネの発色機構
コガネムシの中でも、南米に生息するプラチナコガネは緑や赤といった色調ではなく、金属光沢のような見た目をしています。



図:上からOpima、Betesi、Resplendens
なかなかにピカピカしています。上の赤と緑の虫も含めて、反射スペクトルを測定してみました。
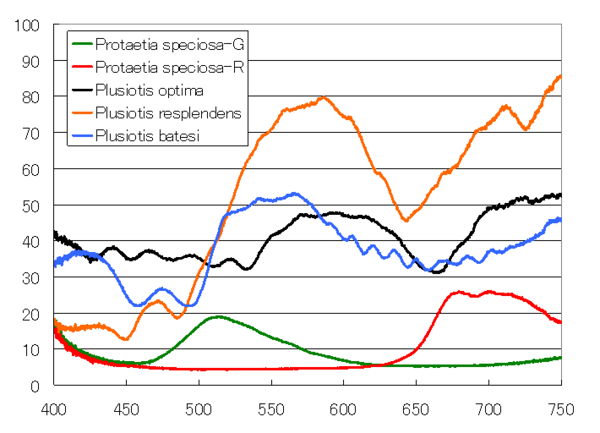
緑と赤の虫は、それぞれの色に対応する波長付近でのみ反射率が高くなっていますが、プラチナコガネは、凸凹はありますが、より広い波長範囲で反射率が高くなっています。プラチナコガネは、どのような機構で普通のコガネムシより広波長範囲で高い反射率を実現しているのでしょうか。周期的な螺線構造による反射のシミュレーションを示しながら説明しようと思います。(シミュレーションは、かなり昔に行ったもので、用いているパラメータが異なる値の方がよい気もしていて、いずれ、異なるパラメータの図にするかもしれません。それにともない、説明の細部は変更されるけれど、筋立ては変更しないと思います。)
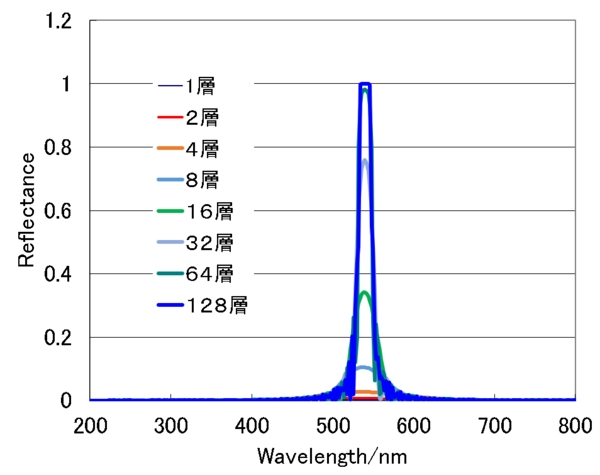
最初のシミュレーションはらせん構造の繰り返し数依存性です。シミュレーションは螺線と同じ符号の円偏光の反射率を計算しています。らせんが1周期(層)では、反射構造は見られません。4周期程度で、少しは反射が生じ、8層、16層と層数が増えるにつれて反射率が上昇し、128層でほぼ飽和状態となっています。
続いてのシミュレーションは、層数を32層に固定して、屈折率の異方性を変化したものです。
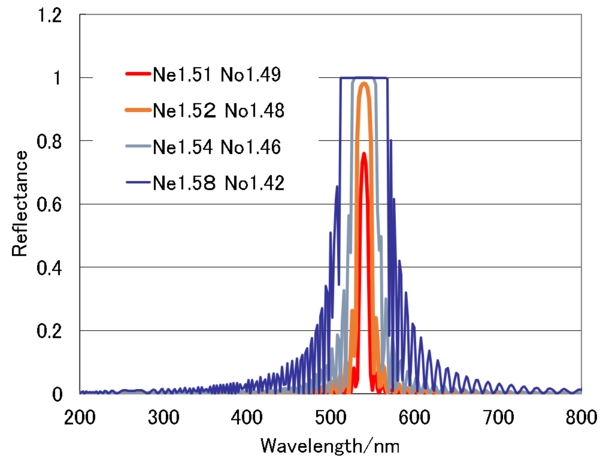
常光屈折率と異常光屈折率の差が0.02のもの(Ne:1.51、No:1.49)は、最初のシミュレーションと同じ条件になります。そこから、屈折率差が広がっていくと層数は同じでも反射率が広がり、そして反射帯の幅も広がっていきます。反射帯の長波長端は、螺線構造の物理的周期をΛとして、NeΛで、単波長端はNoΛなので、NeとNoの差が大きいほど反射帯幅は広くなります。では、400nmから600nmの可視領域全体にわたって反射幅を持つためには、どの程度の屈折率差が必要か計算してみると、Noを1.5とすると、Neは2.25以上である必要がああります。これは、なかなかに大きな値で、これだけ屈折率差のある材料が開発されたら、かなりの話題になるかと思います。プラチナコガネの鞘翅の物質が、このように大きな異方性を有しているとは考えにくいです。
プラチナコガネの広波長反射帯の機構については、1970年代に研究されていて、膜厚方向で螺線周期が変調していることが見出されています[2]。
変調構造を模式的に示したのが上の図です。この図では長周期構造から短周期構造へ滑らかに変化していますが、実際の甲虫では、滑らかというより段階的に変化しているようです。変調構造があると、本当に反射帯が広がるのかを計算してみた結果を示します。
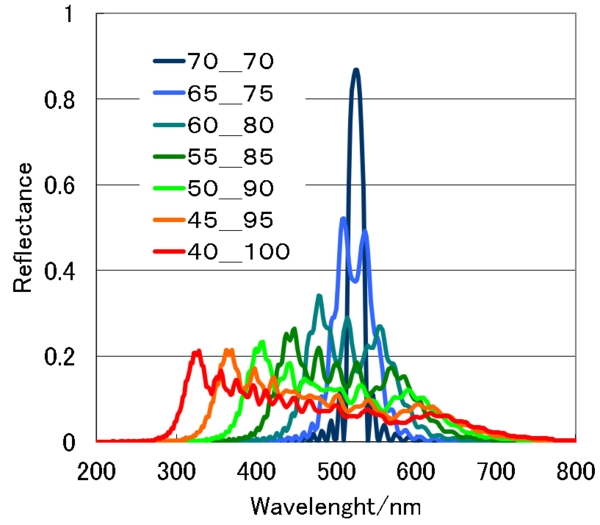
70_70といった表記の数値は、1枚あたり5nmの層が70枚で螺線1周期(350nm)となっているいみです。ですから、40なら、螺線周期は40×5=200nmです。全体の厚さは一定で、その中で、数値で示された範囲の螺線周期が連続して変化するモデルになっています。
見てわかるように螺線周期の変調幅が広がるほど、反射帯の幅も広がります。が、反射率はどんどん低下しており、プラチナコガネの実測値よりはかなり低くなっています。
反射率を上げるには、全体の厚みを増して、層数を増やすか、屈折率異方性を大きくすればよいはずです。試しに、屈折率異方性を大きくしてみたのが下の図です。
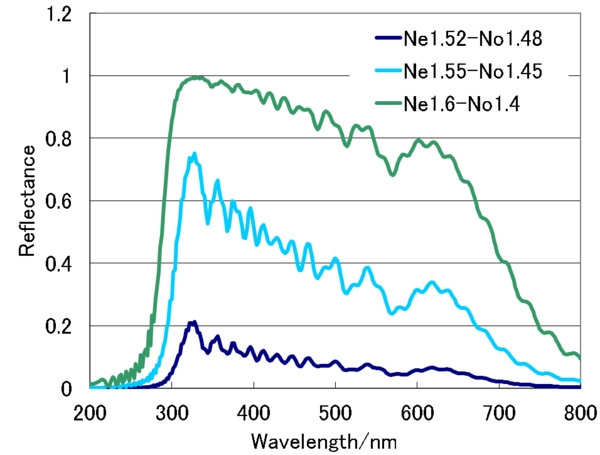
屈折率異方性が0.1あれば、そこそこの反射率に、0.2あれば、ほぼ限界に近い反射率になることが分かります。では、実際の虫の屈折率異方性がどの程度あるかというと……、実測データを見つけたことはないのですが、金属光沢のプラチナコガネについては、鞘翅に複屈折の大きな尿酸を含んでおり、アルカリ性の水溶液で尿酸を洗い出すと、光沢が消えてしまうことが観察されているので、そこそこの異方性を持っているようです。
ところで、螺線構造による反射では、左右円偏光の一方の円偏光しか反射しないので、反射率の最大値は0.5になるはずです。上の計算は入射光を反射する円偏光としているので、最大値が1になっていますが、入射光を直線偏光にすれば(計算では完全な偏光しか扱えないので、偏光していない自然光は取り扱いできません。)、反射率の最大値は0.5になります。改めて、コガネムシの反射スペクトルを見てみると、OpimaとBetesiは反射率の最大値は0.5程度なのですが、Resplendensの最大値は0.8に達しており、一方の円偏光のみを反射しているには大きすぎる値です。
比較のために、BetesiとResplendensを左右円偏光フィルターを通して撮影した写真を次に示します。左が左円偏光、右が右円偏光です。
 
Betesi
 
Resplendens
Betesiは左円偏光では金属光沢を見せますが、右円偏光では金属光沢をうしないます。黒くはなtっていませんが、これは、プラチナコガネは鞘翅にメラニン色素の黒色の部分を持たないためです。話は少しそれますが、深い緑色の虫は鞘翅の奥にメラニン層を持っていて、反射されなかった光をそこで吸収しており、浅い緑色の虫はメラニン層がなく、反射されなかった光がある程度は下部の層で反射されて、明るく浅い緑になっていることが知られています。
さて、Resplendensですが、右円偏光でも鞘翅は金属的光沢を保っています。ただし、色味は左円偏光とは微妙に異なっています。また、足の部分は暗くなっています。少なくとも、足の部分は螺線構造による発色なわけです。
Resplendensの発色機構についても、1970年代に研究が行われています。金属光沢なのは、ほかのプラチナコガネと同様に、螺線周期に変調があるためです。では、右円偏光も反射するのは、左巻きの螺線の間にλ/2波長板を持つ構造によっています。
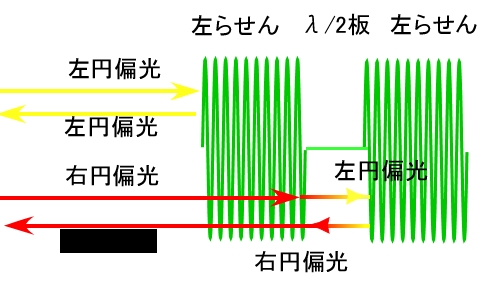
λ/2波長板には、左円偏光を右円偏光に、右円偏光を左円偏光に変換する機能があります。このため、入射光の中の左円偏光成分は、入射後の1層目で反射され、透過した右円偏光はλ/2波長板で左円偏光となり、2層目の螺線構造で反射されます。反射された左円偏光は、λ/2波長板で、右円偏光に変換され、1層目の左螺線で反射されることなく、外部にと出ていきます。
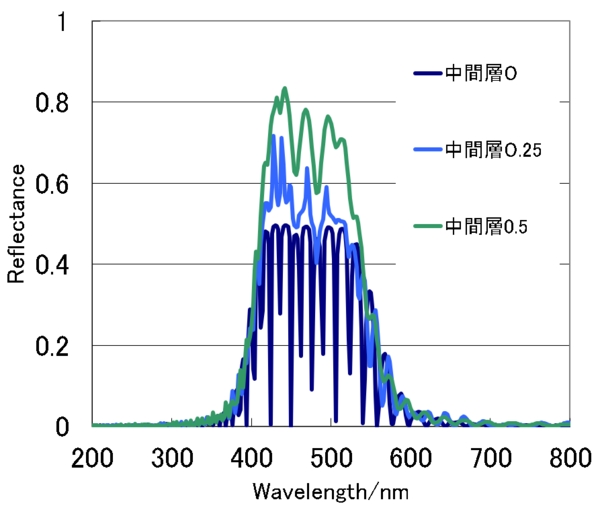
図は中間の位相差板により反射スペクトルがどのようになるかを計算したものです。中間層の厚さが0だと反射率の最大は0.5ですが、波長の0.5倍(λ/2)となると、最大で0.8程度となりResplendensの最大値と同程度となります。中間層が、それ以上に厚くなると、反射率は再び低下していきます。
再帰反射構造を持つコガネムシ
プラチナコガネの一族には、普通の緑色の虫もいます。写真は、キンセスジウグイスコガネで、薄緑色の鞘翅に金属光沢の筋が入っています。金属光沢の筋の部分は、金属光沢のプラチナコガネと同様な広反射帯構造をしているかと思います。

特に注目されていた無視ではなかったのですが、2009年に薄緑に見える部分の構造に関する論文が発表されてから、一部の研究者に興味を持たれるようになりました[3]。薄緑部分を落射顕微鏡で覗いてみた写真を示します。

薄緑の部分は写真のように、基本は六角形の区域に区切られています。六角形の中央は黄色で周辺部は緑で多少青みがかっています。続いて、落射照明の視野絞りを絞って、周辺部には照明光が届かないようにしてみます。
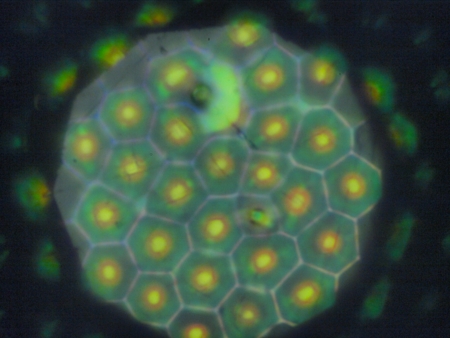
照明光が部分的に当たっている区画を見ると、光が当たっているはずのところは暗く、光が当たっていないはずの領域に色が見えています。六角形の周辺にあたった光は、そのまま戻っているのではなく、内部で反射して区画の反対側で再度反射して戻ってきているのです。
論文には、キンセスジウグイスコガネの六角の区画の構造が、高分子コレステリック液晶の空気界面に見られる欠陥(フォーカルコニック)構造に類似しているとの指摘があり、この研究をうけて、コレステリック液晶で、同様の光学特性を有する膜の作製を目指した研究が、いくつかのグループによって、行われています。
-α版:作業中-
[1] Gold bugs and beyond: a review of iridescence and structural colour mechanisms in beetles, A. E. Seago, P. Brady, J-P Vigneron and T. D. Schultz, J. R. Soc. Interface, Vol. 6, (2009)PPS165
[2] Cuticle reflectivity and optical activity in scarab beetles:
the role of uric acid. S. Caveny , Proc. R. Soc. B V178(1971)pp.205.
[3] Structure Origin of Circularly Polarized Iridescence in
Jweeled Beetles, V. Sharma, et al., Science 325(2009)P449.
|




